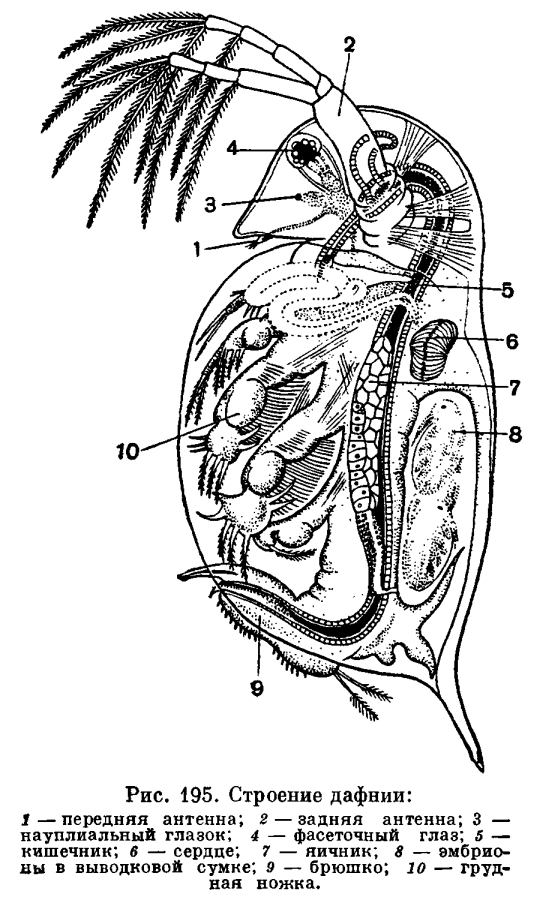
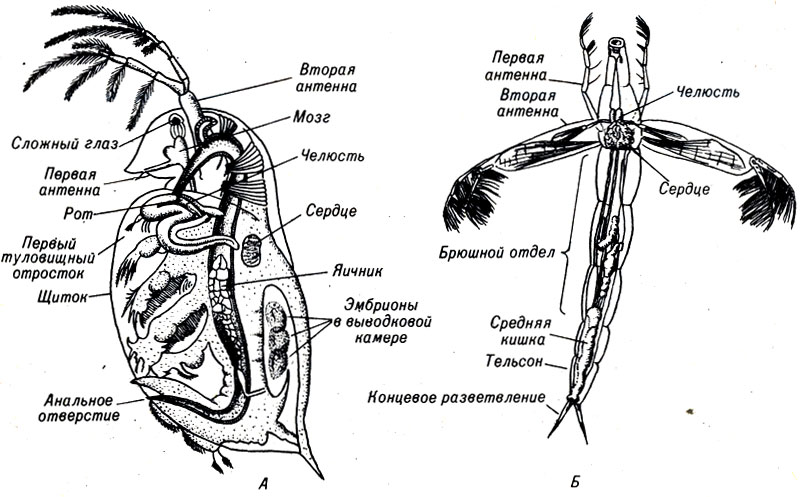
Общие положения
Некоторые объекты, размещенные на сайте, являются интеллектуальной собственностью компании "Живые корма и растения". Использование таких объектов установлено действующим законодательством РФ.
На сайте "Живые корма и растения" имеются ссылки, позволяющие перейти на другие сайты. Компания "Живые корма и растения" не несет ответственности за сведения, публикуемые на этих сайтах и предоставляет ссылки на них только в целях обеспечения удобства для посетителей своего сайта.
Личные сведения и безопасность
Наша компания гарантирует, что никакая полученная от Вас информация никогда и ни при каких условиях не будет предоставлена третьим лицам, за исключением случаев, предусмотренных действующим законодательством Российской Федерации.
В определенных обстоятельствах компания "Живые корма и растения" может попросить Вас зарегистрироваться и предоставить личные сведения. Предоставленная информация используется исключительно в служебных целях, а также для предоставления доступа к специальной информации.
Личные сведения можно изменить, обновить или удалить в любое время в разделе "Аккаунт" > "Профиль".
Чтобы обеспечить Вас информацией определенного рода, компания "Живые корма и растения" с Вашего явного согласия может присылать на указанный при регистрации адрес электронный почты информационные сообщения. В любой момент Вы можете изменить тематику такой рассылки или отказаться от нее.
Как и многие другие сайты, "Живые корма и растения" использует технологию cookie, которая может быть использована для продвижения нашего продукта и измерения эффективности рекламы. Кроме того, с помощь этой технологии "Живые корма и растения" настраивается на работу лично с Вами. В частности без этой технологии невозможна работа с авторизацией в панели управления.
Сведения на данном сайте имеют чисто информативный характер, в них могут быть внесены любые изменения без какого-либо предварительного уведомления.
Чтобы отказаться от дальнейших коммуникаций с нашей компанией, изменить или удалить свою личную информацию, напишите нам через форму обратной связи